Does the Fact that Random Genetic Mutations Can Result in Adaptation Prove Macroevolution?
Image

Michael Behe (Fig. 1), author of Darwin’s Black Box and originator of the concept of “irreducible complexity,” has written his third major book: Darwin Devolves: The New Science About DNA That Challenges Evolution. 1 Behe, a well-known proponent of intelligent design and a senior fellow of the Discovery Institute, 2 is professor of biochemistry at Lehigh University. Over the last three decades, Behe has pointed out many of the unsolved problems for macroevolutionary theory. Behe is a Roman Catholic and can be considered a theistic evolutionist. Behe accepts much of the secular cosmic and biological stories, including descent with modification from common ancestors and deep time. However, based on the evidence in the literature, he rejects the notion that random genetic mutations, acted upon by natural selection, can account for the enormous organized complexity found in the biological world. Instead, he advances the idea that the complexity seen in biology could only have been the work of a supreme intelligence. Although Behe is a theistic evolutionist, his insights into macroevolution are still valuable to the creation/evolution debate. In Darwin Devolves (Fig. 2), Behe argues that although there is evidence that random mutations can lead to increased adaptation of an organism, it does so by breaking things and not by innovation; and innovation is critical to any naturalistic evolutionary theory. Behe says that adaptive variation and evolution of species and genera into other species and genera are possible, but not of families into other families or higher-level taxa. This view amounts to acknowledging variation within biblical kinds while denying unassisted macroevolution. This book review will be written in two parts. Part one (this article) will cover the first six chapters, and part two, due out in February 2020, will cover the rest.
Behe says the “First Rule of Adaptive Evolution” is that evolution can result from the breaking or blunting of any functional gene whose loss would increase the number of species or offspring.
In the introduction to the book, Behe says there are two possible explanations for biological origins: (1) it was an accident, or (2) it was planned. People have held both views for thousands of years. People taking position #1 have pointed to the suffering and evil in nature, suggesting nature could not be the result of a benevolent creator. The design inference dominated western thought from the rise of Christianity until the middle ages when some believed reason and observations alone were better for understanding the world. Paley gave a strong argument for design. 3 Darwin said that natural processes (random variation acted upon by natural selection) working over long time periods could mimic a designer. Alfred Wallace, cofounder with Darwin of the theory of evolution, saw purpose in nature, making him a proponent of intelligent design. Behe suggests that the fine tuning of physics, the lack of contact with intelligent extra-terrestrials, and the discovery of molecular machines all point to intelligent design.
Image

Behe went through graduate school assuming that the molecular details of evolution (abiogenesis and macroevolution) had been worked out and published. Then he read Michael Denton’s Evolution: A Theory in Crisis and came to realize, for the first time, that there were many unsolved problems. He went to the literature looking for evolutionary explanations for the formation of complex molecular machines and only found unsubstantiated “just so stories”! No one had demonstrated how molecular machines could evolve on a biochemical level. Behe became angry. He came to realize that the prevailing acceptance of evolution in academia was more about sociology than experimental science. Behe writes: “When one starts to treat Darwinism as a hypothesis about the biochemical level of life rather than an assumption, it takes about ten minutes to conclude it’s radically inadequate.” Behe sought out and found like-minded scientists with whom he eventually helped establish the Discovery Institute, the flagship organization of the intelligent design movement. His disillusionment with evolutionary theory inspired the writing of Darwin’s Black Box in 1996 followed by The Edge of Evolution in 2007. As science has progressed, the case for intelligent design has deepened. Science has now shown how mutations that are detrimental to biochemical systems can nevertheless be adaptive.
Chapter 1: The Pretense of Knowledge
Behe opens the first chapter with a discussion of brown bears and polar bears. They can mate and bear fertile offspring. Two genes in the polar bear appear to differ from those of the brown bear: APOB, which regulates fat metabolism, and LYST, which controls pigmentation. Apparently, the brown bear “evolved” into the polar bear through damaging mutations in the APOB and LYST genes. So, while the polar bear is better suited for cold and snowy climates than is the brown bear, its adaptations came at the cost of damaged genes.
Behe compares the theory of evolution to theories of economics. Both subjects deal with phenomena involving many variables whose effects are often difficult to quantify. The complexities and uncertainties of these subjects make accurate predictions difficult. Perhaps this is why evolutionary “just so” stories get so little critical push back in academia; an idea only needs to be plausible to get a pass.
Behe identifies three red flags concerning evolutionary theory. First, the literature is full of “just so” stories that have little or no evidentiary backing. Thus, there are lots of stories with little evidence; Behe says this situation amounts to a pretense of knowledge since most of what passes for knowledge is mere untested speculation. Behe calls the second red flag the “united front.” Many evolutionary biologists today show a united front against contradictory evidence such as found in developmental biology and epigenetics, yet many privately admit Darwinism is deficient. The third red flag arises from what Behe calls the “Principle of Comparative Difficulty.” This principle states that if a relatively simple subject is riddled with uncertainties making it hard to understand, a relatively more complex subject will be even more difficult to understand. As a relatively simple subject, Behe refers to human dietary practices and their effects on health. For example, much of what was once held true about the consumption of red meat, cholesterol, eggs, coffee, fat, salt, etc. has been overturned by more study. It took years of research in real time on thousands of people to reach the correct understandings. How much more difficult is it to explain the evolution of life, which can’t be studied in real time and is orders of magnitude more complex. Behe’s point is this: if scientists working in the here-and-now have difficulty understanding how health is impacted by eating habits, how much more difficult is attempting to understand evolutionary processes that can’t be directly observed?
Complex systems controlled by myriad variables are unpredictable, so there is a limit to knowledge of these systems. Behe considers evolution to be unpredictable; we can’t ever really understand the causes of life’s history, nor even accurately determine its history. So, any “just so” stories triumphantly declaring how evolution worked are mere pretenses of knowledge.
Behe says there are three levels of explanation for phenomena. There are things we can know, things we can’t know, and things we know have an intelligent cause. Causes we can know may be perceived through direct observation or perhaps strong statistical correlation. When there are only weak correlations between cause and effect in complex systems, we can’t be certain how the variables bring about the effect. Behe says that in those cases the cause is chaos. Long term weather forecasting and evolutionary narratives fit this case. Behe challenges us to imagine the difficulty of accurately modeling the weather on earth over the last several millennia while considering the difficulty we have predicting the weather for the next month. And if that is difficult to get right, how much more difficult is it to model the past evolution of life on earth over billions of years?! Intelligent causes are distinct, being able to produce complex specified information and operate independently from physical causes. Behe closes the chapter by asking if it is possible to at least find out what level of explanation best fits the appearance of life on earth.
Chapter 2: Fathomless Elegance
In this chapter, Behe provides a flavor of what an explanation for life must account for. He briefly describes what modern technology has enabled us to learn about living things such as the reactions and structure of DNA and proteins; the structure, construction, and function of molecular machines; how cells behave; etc. He then turns to specific examples.
Planthoppers are insects that can jump hundreds of times their body length. In order to do this, their hind legs must move simultaneously. It turns out that the hind legs are propelled by gears that spin at 50,000 teeth per second! 4 The gears facilitate coordination of movement.
Behe then turns to the vertebrate camera eye. Darwin thought the human eye could evolve by the variation/natural selection mechanism, but he had no knowledge of the complexity at the molecular level. Modern Darwinists claim the eye is flawed, so it could not have been designed. But they fail on two counts. One, logically, even an object with flaws could have been designed. Second, it turns out that the perception of flaws was due to our ignorance of the design! 5 It was thought that the eye was wired backwards and that the blind spot limited vision. But subsequent research has revealed cells that act like optical fibers embedded in the retina. They deliver all the optical information to the rods and cones (sensory cells) despite the latter being located far from the source of light. It turns out that the blind spot of one eye is covered by the scope of vision of the other eye, so there are no gaps in the field of view.
There are birds which navigate by the magnetic field of the earth. There are bacteria that do the same. They are known as magnetotactic bacteria. The cells make little iron crystals that act as magnets. The particles must be fashioned into the right shape and size in a compartment separated from the rest of the cell due to the iron’s toxicity. This process requires specific proteins. The ratio of Fe(II)/Fe(III) must be tightly controlled. The numerous iron crystals in their separate compartments must then be positioned and aligned, resulting in a net large magnet. The coordination of this activity is on the scale of a human manufacturing facility. The production and alignment of the magnetic particles involves 12 genes. According to Behe, “Experimental disruption of any of a dozen different genes results in either the severe weakening or complete elimination of the cell’s magnetic response,” suggesting the system is irreducibly complex.
Several means of locomotion for bacteria are described. There are bacteria that have treads like tanks and secrete a polysaccharide road for traction, some have rotary engines, some have leg-like projections, and some have an internal flagellum that gives the cell a cork-screw appearance and by spinning propels the cell forward. All of these means of locomotion involve many parts and types of proteins and the attendant regulatory networks that control their construction.
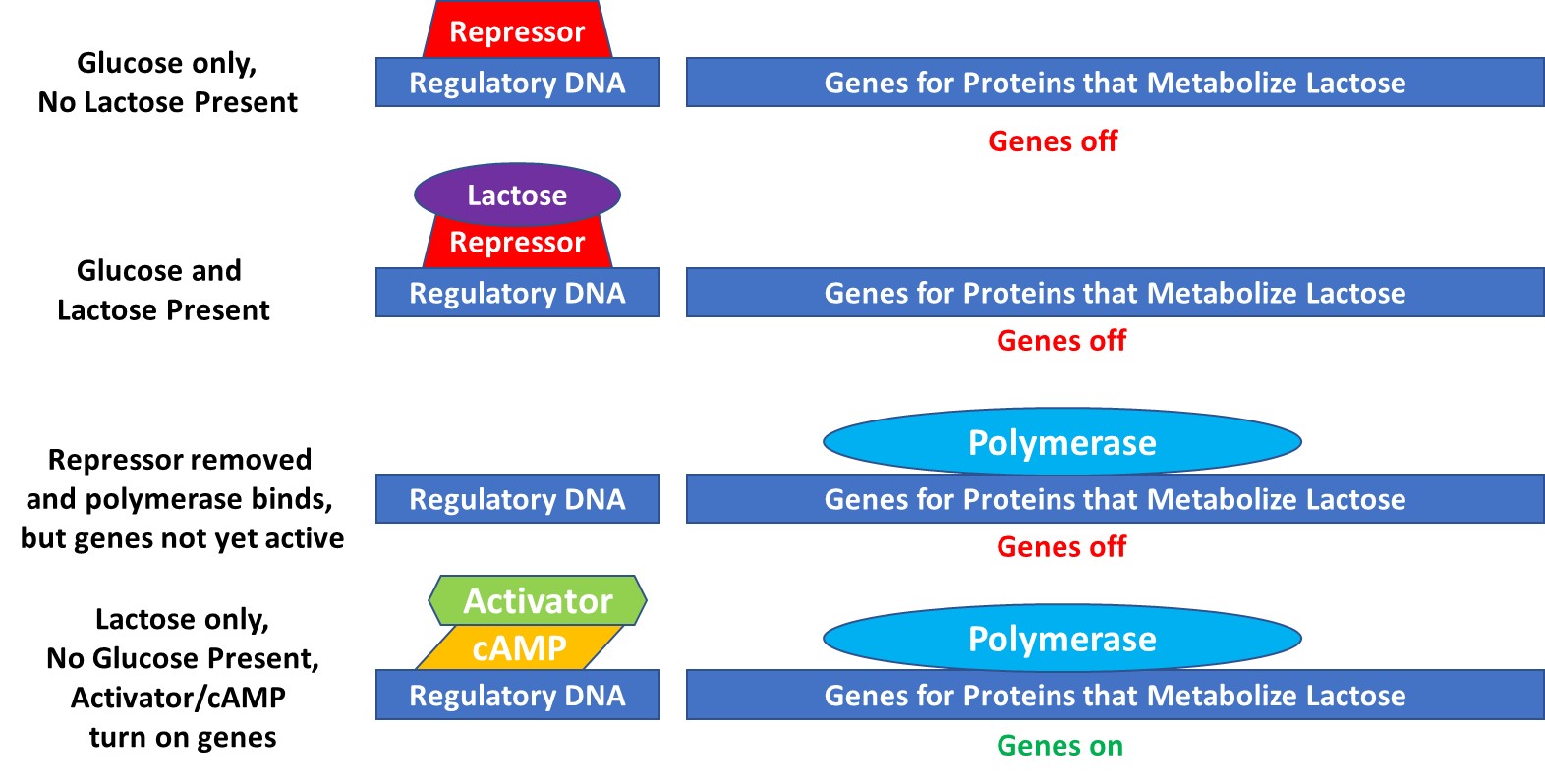
Behe describes in some detail how E. coli controls when and where it can metabolize lactose instead of glucose. E. coli can eat either sugar when present exclusively, but prefers glucose in the presence of both. The control mechanism for determining which sugar to metabolize is complex and involves regulatory DNA, repressors, activators, polymerases, ribosomes, genes for metabolizing proteins, m-RNA, and more. In Figure 3, the sequence of regulatory events leading to the activation of the genes that code for proteins that metabolize lactose is shown.
The genes that code for the proteins for lactose metabolism are controlled by nearby regulatory DNA. In the absence of lactose, a repressor molecule is bound to the regulatory site. In the presence of both glucose and lactose, lactose binds to the repressor molecule, thereby changing its shape, resulting in its detachment from the regulatory DNA. The absence of the repressor molecule facilitates placement of the polymerase (a molecular complex that copies DNA into m-RNA) on the genes, but the genes are not yet activated. Then, in the absence of glucose, an activator molecule combined with cAMP (which only appears in the absence of glucose) binds to the regulatory DNA, thereby activating the genes and initiating m-RNA production. This level of control is similar to computer programming.
Behe next discusses alternative splicing. It has been long understood that DNA is transcribed into m-RNA that is then translated into proteins in ribosomes. However, we now know that a single gene can be transcribed into potentially thousands of m-RNAs by a molecular machine known as the spliceosome. The original m-RNA transcript can be edited by the removal of selected pieces and then stitched back together. The final m-RNA is then sent to the ribosome for translation into a protein. The specific editing is determined by the spliceosome. The spliceosome consists of several RNA molecules plus hundreds of different proteins. The spliceosome is dynamic and programmable with various proteins coming and going as needed. The proteins help determine where and when to splice.
In eukaryotic cells, regulatory DNA may not be close to the gene it controls. Eukaryotic genes may have dozens of control factors (molecules that can bind to the regulatory DNA). The distance between the regulatory DNA and the gene it controls can be shortened by scaffolding proteins that bring the two DNA strands together. The orchestration of bringing together genes and their regulatory DNA is a hot research topic.
Chapter 3: Synthesizing Evolution
Recall that Behe is a theistic evolutionist. He opens this chapter with what he considers evidences for common descent, the part of evolutionary theory he accepts. 6 He cites the fossil record: the same types of fossils are found in the same strata in different geographic locations. Similar traits among organisms suggests common ancestry. Similarity of traits and proximity go hand in hand and suggest common ancestry. Darwin claimed that if special creation were true, organisms with similar traits would be found in separate localities with similar climates; similarities would not be correlated with localities. The field of biogeography examines the distribution of plants and animals geographically in an attempt to understand evolutionary history.
Behe recites the theory of how the continents formed. He said that a supercontinent, Pangea, broke up. First Australia separated, then Africa, then South America, and finally North America and Europe. This order of events is consistent with similarities of organisms across continents.
Behe briefly describes how genetic inheritance works. He discusses Mendel’s pea experiments. We eventually learned that DNA carries the information of inheritance.
Behe mentions that phylogenetic trees based on morphology and those based on molecules don’t always agree. He says comparing molecules does not give clear phylogenies due to horizontal gene transfer among bacteria. Genetic mutations are usually the result of copying errors during reproduction and recombination.
Behe describes microevolution as something Darwinian processes can accomplish—evolution of a species or genus into a new species or genus. Macroevolution involves evolution of higher taxa. Behe says most scientists accept common descent, but many question the role of random mutations.
Darwin rejected a creator because he considered aspects of nature as cruel with much suffering, something he believed a benevolent creator would not allow. Hence random variation without purpose or direction became the default creative agent. Behe wants to examine if random variation can support the explanatory weight placed upon it.
Chapter 4: Magic Numbers
Behe surveys the latest thinking on Neo-Darwinism. Realizing the weaknesses of Neo-Darwinism, many biologists have sought alternative explanations. First, Behe considers “Neutral Theory,” which says that small changes to proteins don’t affect survival, so are invisible to natural selection. Behe doubts all non-coding DNA has function, contrary to most young-earth creationists and even other intelligent-design advocates. Specific neutral mutations have a poor chance of being fixed in large populations. Nevertheless, according to the theory, a small, steady number of new, neutral mutations becomes fixed in a population every generation, regardless of population size. The number is similar to the number of new, neutral mutations in each offspring. For humans, this number is 10 to 100 new mutations. This generational mutation rate is what is used to set molecular clocks.
Most DNA in bacteria is coding, but in humans most is non-coding. The evolution of eukaryotic cells is thought to have resulted from gene duplication, leading to larger genomes. Neutral duplicated genes are unlikely to be fixed except in small populations. But just duplicating a gene does not account for the information in it or the new information required for evolution. The source of variation is still left to chance. The source of the required beneficial mutations to drive macroevolution is the unanswered question.
On a genetic level, life does not look like a tree but like a web. Horizontal gene transfer may help explain some of the pattern for microbes. Genes in microbes appear to be randomly distributed without evidence of linear ancestor-descendant relationships. Some scientists think that after abiogenesis, the genes were duplicated and mutated into everything else through genetic drift (neutral evolution) and weak natural selection.
Next Behe discusses the origin of life and the multiverse. An infinite number of universes will make inevitable even very unlikely events, including the origin of life. Koonin calculated the probability for abiogenesis in our universe is 10-1018! But with an infinite number of universes, anything can happen. This approach attempts to explain biological unknowns with cosmological speculations backed by zero evidence; Las Vegas anyone?!
Complexity theory is considered next. Attempts are made to model complex dynamic systems with computers. There are examples of self-organization in nature: hurricanes, schools of fish, flocks of birds, crystal formation, etc. The thinking is that perhaps complex systems emerge spontaneously. In a computer program called the Game of Life, only a very small subset of rules and possible initial conditions provided the potential for evolving dynamic complex systems. Everything else resulted in very simple systems unable to carry information. Complexity theory has yet to shed light on how life could have evolved.
Behe concludes this chapter by observing: “Breathing the thin air of abstract theory for too long can induce hallucinations, and we start to imagine phantasms transforming themselves into whatever we wish to see.”
Chapter 5: Overextended
Chapter 5 deals with what has been dubbed the “Extended Evolutionary Synthesis.” Instead of abandoning Neo-Darwinism, some have sought to patch it up. One such attempt is referred to as Evo-Devo, which is short for Evolutionary Developmental Biology. Evo-Devo studies how a single fertilized cell develops into an adult form. Development is orchestrated by master genes and proteins. Many master genes are similar across a wide assortment of organisms, presumably because of common ancestry. For example, the master gene PAX6 regulates eye development in flies and vertebrates. Master genes activate cascades of events leading to the development of the embryo. Master genes are like switches that activate other genetic programming. The proteins produced by master genes bind regulatory DNA sequences near the genes they activate. Theorists speculate that evolution was driven by mutations in the regions near genes. A mutated region might then bind a protein generated by a master gene and come under its control. This theory provides no explanation for the origin of master genes or developmental regulatory networks, nor are any detailed explanations for the origins of switches and connections made. Behe asks what master genes would need to change to produce the gears in planthoppers? Three decades after the discovery of master genes, there are no detailed accounts of how changing them could account for some complex structure.
Behe next discusses inclusive inheritance. Inclusive inheritance considers the role of epigenetic factors in evolution. Epigenetic factors are heritable features other than the sequence of DNA. For example, DNA can be tagged with methyl and acetyl groups, affecting the expression of genes without changing the DNA sequence. Some think variation in epigenetic information could give evolution something to work with. Some think combinations of changes in epigenetics could result in some benefit. However, multiple simultaneous epigenetic changes are likely to be detrimental.
Behe next discusses niche construction. The idea here is that as organisms modify their environment, the new environment will shape future offspring. Hence, this view sees organisms as driving their own evolution. While modifications to the environment could be an impetus to adaptation, it does not explain how that pressure is translated into new complex structures in an evolving organism.
Behe then turns to “Developmental Plasticity,” which refers to the capacity of an organism to change its phenotype in order to adapt to its environment. There is a range of variation in the phenotype of an organism that can occur without changing the DNA. According to this theory, environmental stress could result in a change in phenotype. With the better phenotype in place, the right mutations could then fix the altered phenotype as the new norm. This theory is speculative and still needs to explain where the right beneficial mutations come from.
Next considered is “Natural Genetic Engineering.” This theory assumes complex systems are already in place and does not attempt to explain their origin. According to this theory, the molecular tools used for modifying DNA can somehow respond to selection pressures by processing DNA to make it more adaptive. Hence cells can use their DNA editing capabilities to guide their own evolution. Behe says there are limits to what cellular processes can adapt to. If this idea proves true, it would suggest a built-in genetic mechanism for adaptation pointing to design.
There are allegedly eight major transitions in evolutionary history, the first being the origin of life. So far, there is no credible account for how chemistry could have become biochemistry. All experiments to date have barely scratched the surface, if that. The remaining seven transitions are purely speculative.
Many theories explore the necessary conditions for the evolution of life without even considering how things could happen on a molecular level. And evolution at the molecular level, of course, is what needs to be explained!
Chapter 6: The Family Line
In chapter 6, Behe provides evidence for microevolution, but not macroevolution. Detailed investigations of the history of mutations in specific populations have only been going on over the last twenty years. Only now are we finally able to begin to test Neo-Darwinism. Do random mutations acted upon by natural selection lead to new complex functional features? New results suggest strict limits to change. The observed changes can account for evolution of one species into a new species and one genus into a new genus, but not evolution of higher classification levels.
Darwin found between twelve and fourteen species of finches on the Galapagos Islands. The species varied in beak sizes and shapes. The large beaks were good for cracking tough seeds while the smaller and probing beaks were good for getting nectar. Peter and Rosemary Grant have been studying the Galapagos finches over the last several decades. 7 They make a yearly trip to the islands. They examine the birds under a wide variety of circumstances. There are variable climates on the islands. The islands are of volcanic origin so have different climates at various altitudes. During periods of drought, the larger birds with larger beaks fared better because they were able to crack into the prevailing tough large seeds. After the drought, the average size of the surviving birds had increased in size by five percent. However, all this reversed when the rains returned.
According to Behe, the finches migrated to the islands 2 million years ago. Since that time, there has been little change. The finches have remained finches.
Recently, 120 Galapagos finches had their genomes sequenced. Two mutations were found in the ALX1 gene. The ALX1 gene codes for a protein that controls beak shape and size: blunt or sharp. Both mutants have been on the islands since the first migration. Climate determines the gene frequencies. The mutations are thought to damage the gene for sharp beaks resulting in blunt beaks. Hence the mutations lead to adaptation by damaging a gene. The observed variations were at the genus/species level only; no higher taxa were involved. There are many varieties of life on the islands, so the possible mutations were not restricted by the environment. Behe asks if two million years is long enough to test the Neo-Darwinian mechanism. He points out that the Cambrian Explosion mostly took place over ten million years when scores of new phyla were formed. Behe says that whales presumably evolved from a terrestrial ancestor over a ten-million-year period. Behe says that the finch data demonstrate that the “edge of evolution” is between the genus and family level; Neo-Darwinism can account for microevolution of species and genera but no further. The origin of families and higher taxa requires design.
Behe says that descent with modification from a common ancestor and purposeful design are not in conflict. I think most creationists would agree with this statement although they would take its meaning differently; God created all living things after their kinds with built-in programming for variation for adaptation within limits. Darwinists need to demonstrate that chance variation can create novel biological information.
Behe discusses various other disparate living things. He examines how many species, genera, families, and high taxa have evolved from a small initial population having opportunity to evolve. In every case the result was the same: even in environments with variable climates, the level of variation was always at the species/genus level and not higher. The various life forms were in environments that gave evolution opportunity to shine. Many were on islands or in recently filled lakes. According to Behe, there was time to diversify. There were only a few species initially. And yet, in example after example, variations were limited to a species and a few genera and no more (see Table 1). 8
Table 1: New Classifications Produced by Luxuriantly Evolving Groups
| Species | Genera | Families | Higher Classifications | |
|---|---|---|---|---|
| Finches | 14 | 4 | 0 | 0 |
| Cichlids | ~1500 | ~75 | 0 | 0 |
| Anoles | ~300 | 3 | 0 | 0 |
| Honeycreepers | 55 | 24 | 0 | 0 |
| Fruit Flies | 1000 | 2 | 0 | 0 |
| Beetles | 239 | 1 | 0 | 0 |
| Silverswords | 50 | 3 | 0 | 0 |
| Lobelias | 126 | 6 | 0 | 0 |
| The Family Line | ||||
Relatives to the above life forms can be found in other parts of the world; all belong to the same families.
Well, that’s all this month. In part 2, due out in February, we’ll cover the rest of the book.
- 1Behe MJ (2019) Darwin Devolves, Harper One, New York, NY
- 2Discovery Institute, Michael J. Behe. https://www.discovery.org/p/behe/ Accessed 2019 Dec 21
- 3Coppedge, D (2018 Mar 30) Paley’s Watch Found. https://crev.info/2018/03/paleys-watch-found/ Acessed 2019 Dec 21
- 4Behe provides a photograph of the gears on p. 46. You can see the picture online. Look up the book on Amazon, click on the image of the book cover, then advance to page 46. There is also a video: www.youtube.com/watch?v=xQk-lP2R04Y
- 5See https://tasc-creationscience.org/article/review-jonathan-wells%E2%80%99s-newbook-zombie-science-part-2 for a discussion of the human eye and its alleged flaws.
- 6Young earth creationists accept microevolution but not macroevolution. Behe accepts common ancestry for all of life, young earth creationists accept common ancestry within created kinds only. Similar organisms lived in similar habitats and hence were buried in similar layers during the Flood. Organisms have common traits because they had a common designer. The global Flood and the repopulation of the world with terrestrial organisms explains the data of biogeography and does not contradict special creation.
- 7Peter and Rosemary Grant have pioneered a detailed and in-depth study of finch evolution in the Galapagos over the last several decades. The have published many books and articles on the subject including their Pulitzer Prize winning book The Beak of the Finch.
- 8Behe MJ, Table 6.3, 167.