The Ear: Evolved or Designed? - Part 1
Previously I wrote an article about how the complexity of the human ear implies a designer rather than evolutionary origin. Evolutionists attempt to explain the existence of such complex organs by gradual changes over time. In this article I would like to explore the diversity of hearing organs in different creatures and discuss whether there is evidence of gradual changes in different organisms which would lend credence to evolutionary theory or if the diversity of ears is evidence for the improbability of evolution and evidence of design by a Creator. Before we look at the different ears, I would like to define a few scientific concepts which come into play when discussing evolution: homologous, convergent, and divergent evolution and phylogenetic origins.
In phylogenetics the characteristics of different organisms are used to group organisms into families and create a tree that is assumed to demonstrate their origins. Organisms with similar characteristics are assumed to be more closely related. The similar structures are thought to be evidence for their close relationship. The phylogenetic tree (Fig. 1) is basically the family tree that evolutionists have created to show the theoretical relationships between different organisms and how they all evolved from some single-celled organism over billions of years. It is organized based on the assumption that similar characteristics or similar DNA imply a close relationship.
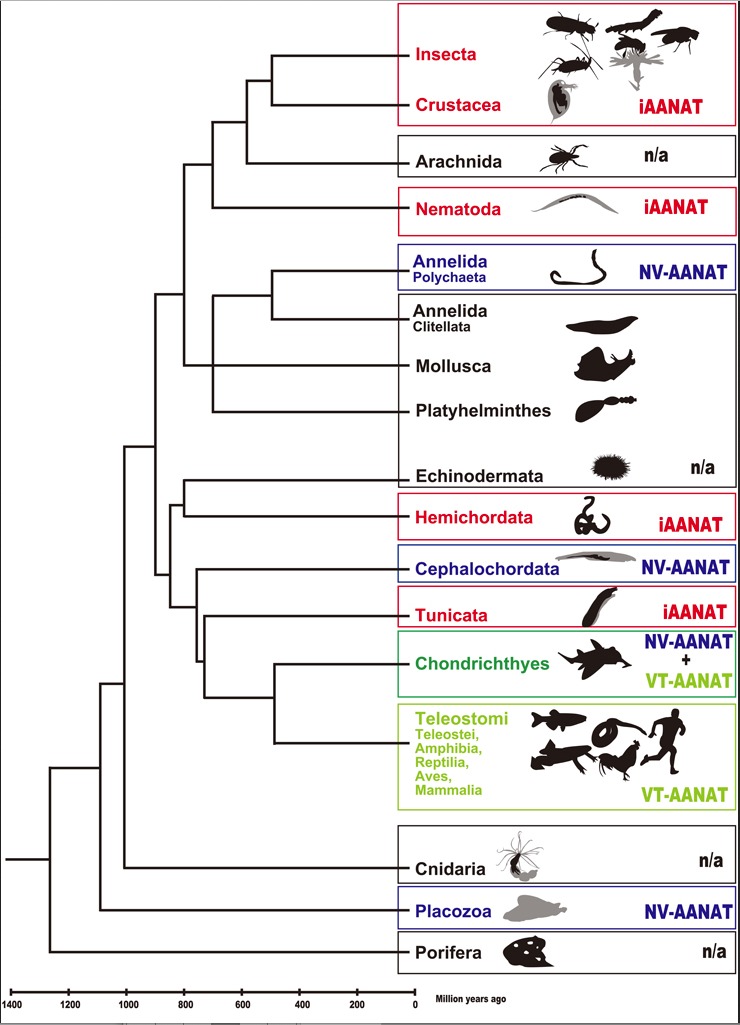
Figure 1 Schematic phylogenetic time tree. HiragakiS, Suzuki T, Mohamed AAM, Takeda M. Structures and functions of insect arylalkylamine N-acetyltransferase (iaaNAT); a key enzyme for physiological and behavioral switch in arthropods. Front. Physiol. 6:113. doi: 10.3389/fphys.2015.00113
Homologous structures are structures that have a similar appearance, but perform different functions. Evolutionists theorize that one structure evolved into another. Homologous structures are thought to be evidence for evolution from one organism into another organism.
Analogous structures exist that perform similar function, but have no common ancestor. These structures are considered evidence of convergent evolution.
In Figure 2, the cat leg and the whale flipper are thought to be homologous because the whale is thought to be evolved from a mammal. Since the cat and the praying mantis are not thought to be related, their legs are said to be analogous rather than homologous.
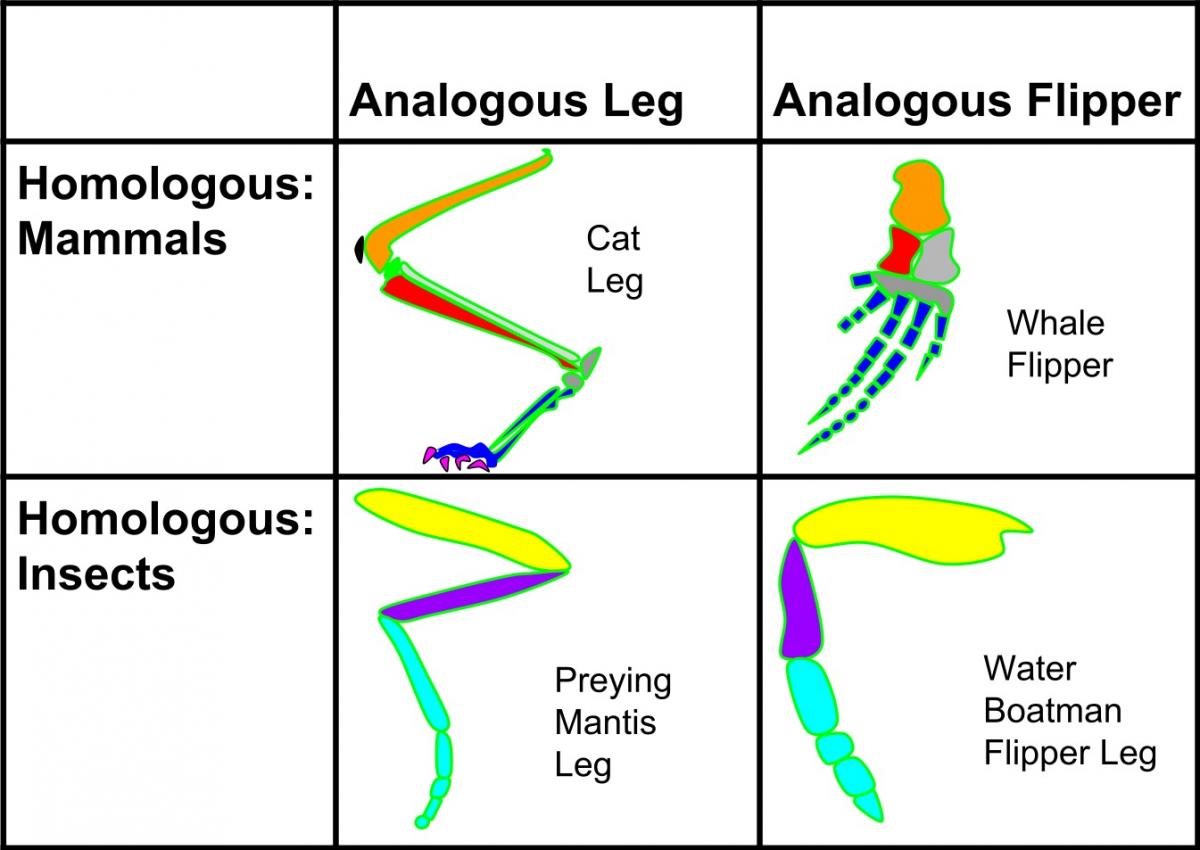
Figure 2 - By Vanessablakegraham - Own work, CC BY-SA 4.0, https://commons.wikimedia.org/w/index.php?curid=49147917.
The difference between homologous and convergent evolution is in the supposed phylogenetic origin of the structures.
If two organisms which are thought to be closely related have a similar characteristic, they are called homologous and the similarities that they share are considered as evidence for their evolution. So creatures which are closely related would have homologous structures or homologous DNA because that is how they were determined to be related. Now when a creature has a characteristic or structure or DNA which is like that of another creature, but it has been assumed that they are not closely related, then these structures are called analogous and are said to be evidence of convergent evolution. So even though they have similar structures or similar DNA, they are not considered evidence for a close relationship because they have been predetermined to be unrelated.
Evolutionists use of homologous structures is a good example of circular reasoning. Circular reasoning is when the conclusion is assumed to be true and used as the evidence to prove itself. So, homologous structures are what suggest the phylogenetic tree to the evolutionist and are also what is considered evidence to prove that evolution is true, except in the case when the organisms are not thought to be related. Then the similar characteristics are not used to prove that they evolved from one another. Rather, the similar structures are said to have evolved convergently. Since evolution is assumed to be true, then it is thought that these characteristics evolved multiple times. It is assumed that their environments selected for these traits. Yet natural selection can only select from genes that have randomly mutated. It is accepted by both evolutionists and creationists that environmental factors select for certain traits, but these traits have to come into existence. Selection pressure cannot act on the mutations to cause a desired genetic sequence or trait to arise. Evolutionists have no explanation for how the environment could influence random mutations which create similar traits. In defining these terms without close examination, they appear to be evidence for evolution, but in the case of a highly specialized function which appears in multiple places on the assumed phylogenetic tree, they are further evidence of the improbability of evolution.
In an article in Evolution News, Jonathan M. [last name not provided] puts it this way:
The methodology for inferring common descent is thus dependent on circular reasoning. If similarity can be accounted for with reference to common descent, then it is taken as evidence for common descent. On the other hand, if similarity cannot be accounted for with reference to common descent, it is evidence for convergent evolution. It’s a typical “heads-I-win, tails-you-lose” tactic.
The methodology is circular—it assumes that these structures are connected by descent. When one’s entire interpretative framework presupposes common ancestry at the outset, it is no wonder that any and every observation is taken as supportive of that paradigm. 1
An example of this is echolocation. Bats and dolphins can transmit high-frequency sounds, their ears are specialized to hear the reflections of these sounds and their brains are equipped to “see” their surroundings like we see a baby using ultrasound. According to the accepted phylogenetic tree, bats and dolphins are not thought to be closely related, so echolocation must have evolved multiple times.
Random genetic mutations evolving this remarkable capability once would seem beyond the realm of probability. To think that this could have evolved separately in two unrelated species defies reason.
A study published in Nature in 2013, comparing gene sequences associated with hearing in bats and dolphins found similarities:
Strong and significant support for convergence among bats and the bottlenose dolphin was seen in numerous genes linked to hearing or deafness, consistent with an involvement in echolocation. Unexpectedly, we also found convergence in many genes linked to vision: the convergent signal of many sensory genes was robustly correlated with the strength of natural selection. This first attempt to detect genome-wide convergent sequence evolution across divergent taxa reveals the phenomenon to be much more pervasive than previously recognized. 2
They were surprised to find that bats and dolphins had many similar gene sequences related to vision and hearing even though they are not thought to be closely related. This would seem to indicate that these gene sequences occurred randomly multiple times, a miracle even less probable than winning the lottery multiple times. They attribute these similarities to convergent evolution.
Evolutionists use natural selection as a mechanism to account for this improbable occurrence, but they neglect the fact that natural selection can only act on mutations which have occurred randomly. Natural selection does not provide an explanation for random mutations resulting in the same genetic sequence more than once. The statement below is an example of this line of thinking.
Hearing has independently evolved multiple times, thus our particular design of hearing is clearly very important. It is rare for random mutations to appear, however, if they do appear and stick around multiple times, this must mean that a very strong selection pressure exerted on them. 3
Homology has primarily been observed phenotypically, in the visible characteristics, but it is assumed that these structures are passed from one species to another through their genetic code. It is expected that the genetic code of closely related organisms would be similar, but for an unrelated organism to develop analogous structures using similar genetic code is highly unlikely to have occurred randomly and speaks to the signature of a designer.
Since we have just recently been able to decipher the genetic code and still don’t understand how it causes organ development, evolutionists have looked to embryological development for evidence of inheritance. It is assumed that similar genes would be inherited by evolving species, and these genes would coordinate similar development. Noted evolutionist, Gavin de Beers, wrote in his treatise, Homology, an Unsolved Problem, that structures that were thought to be homologous were arising from different portions of the embryo which would lead one to believe that they had a different genetic cause and thus were not inherited traits and had not evolved from one another. 4 If similar structures arising on different parts of the body are a problem for evolutionists, then insect ears would seem to be a real problem.
Insect Ears
Insect ears can be found at almost any location on an insect’s body. Figure 3a presents a diagram of Drosophila showing several body locations where ears have been identified. According to David Yager, who wrote a literature review article on insect auditory systems, ears have appeared independently at least 19 different times in the Class Insecta. 5
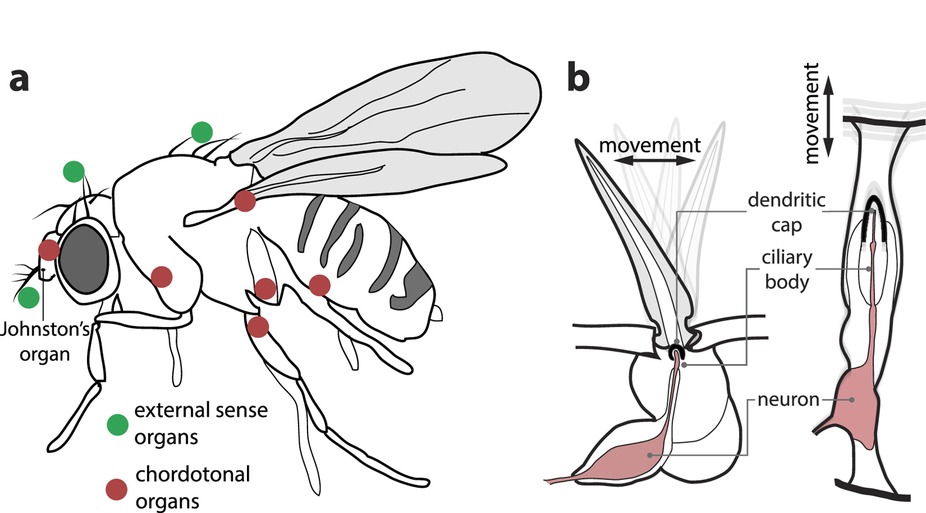
Figure 3 -
Sensory hair cells and chordotonal organs.
(a) Locations of sensory hair cells (including the antennal receiver) and chordotonal organs in Drosophila.
(b) Insect arista and chordotonal organs.
Lorimer T, Gomez F, Stoop R (2015) Mammalian cochlea as a physics guided evolution-optimized hearing sensor. Sci Rep 5:12492
How do evolutionists explain this diversity? This is the explanation given in a post for What-When-How.com:
We now know through developmental studies and comparative anatomy that it is not so difficult to “construct” an ear by making a few peripheral modifications to an existing proprioceptor and its surrounding cuticular and tracheal structures. By simply thinning the cuticle, enlarging the surrounding tracheal air sacs to allow membrane vibration, and mechanically isolating the sensory organ from body movements, a proprioceptor can be converted into a sound-pressure receiver (Fig. 4). For example, the bat-detecting ears on the thorax of noctuoid moths are thought to have evolved from proprioceptors monitoring wing movements. 6
Evolutionists often consider body parts like boards on a construction site. One that is no longer needed can easily be cut down to size and used for a different purpose. Totally discounting the fact that this requires a designer and that in life these changes would be orchestrated by genetic control. It is like making a change in the software for your laptop that allows it to create a new laptop with a lighted keyboard or touch-screen. This is such a simple modification, but it certainly didn’t happen randomly. Even if random genetic changes could convert wing-monitoring proprioceptors into ears, these changes would have to happen concurrently with changes in the insect’s central nervous system that would allow for new interpretation of the information. Otherwise, whenever a potential mate would fly by, instead of hearing the sound and flying after her, his brain would interpret the sound as the movement of his own wings and think that he was flying. He would have no idea where he was going because he would have no clue he should be chasing after the sound that he thinks is the movement of his wings.
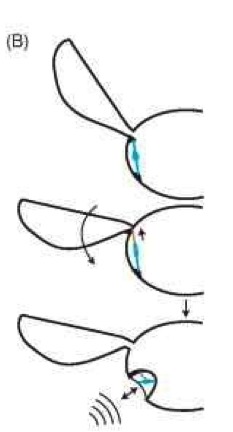
Figure 4
Not only are insect ears found in many different locations on the insect’s body. There are many different types of insect ears. All ears must have some form of receptor for sound and a mechanism to convert the movement of the receptor into nerve transmission to the brain. Two types of insect ears are tympanal ears and antennal ears. In an antennal ear the antenna directly and mechanically causes the opening of channels on the nerve cells instigating an electrical signal to the brain. In a tympanal ear the tympanum stimulates movement, directly or indirectly of fluid and the movement of the fluid opens the channels on the nerve cells.
Antennal Ears
Female fruit flies use antennal ears to hear the call of their prospective mates. As part of the fruit fly mating behavior, males vibrate their wings (instead of rubbing their wings together) making a specific mating call which varies according to species. The female hears the mating call through a specialized antenna called a Johnston’s organ (Fig. 3b), which is tuned so that the best frequency correlates with the song of the males of her species. The antenna of the Johnston’s organ vibrates in response to the male’s wing movement. 7
The frequency of this sound is between 100 and 300 Hz, which is within human hearing range, but it is too quiet for us to hear. 8 The movement of the antenna mechanically opens gated channels in nerve cells directly creating a nerve impulse which is sent to the brain. 9
Tympanal Ears
Mammalian ears have a tympanic membrane which vibrates causing movement of the middle ear bones or ossicles which act as a lever system to stimulate fluid movement inside the cochlea (Fig. 5). The bones act as an amplifier which overcomes the barrier to sound which occurs at the air to liquid interface. A katydid has a similar system; however, the tympanum of the katydid does not push on the lever; it is one end of the lever. As it vibrates it causes movement of fluid in a chordotonal organ. The fluid then causes movement of the nerve cell receptors. This simple system is very efficient and allows the katydid a broad hearing range of 10,000 to 50,000 Hz. This is important because the mating call of the katydid is around 23,000 Hz and it may also allow the katydid to hear bats before becoming their prey. 10 For comparison purposes humans hear from 20 to 20,000 Hz. The human cochlea is rolled up like a snail and is about 3 cm long. The katydid’s acoustic vesicle is 600 microns long and could only be seen by using microscopic CAT scans.
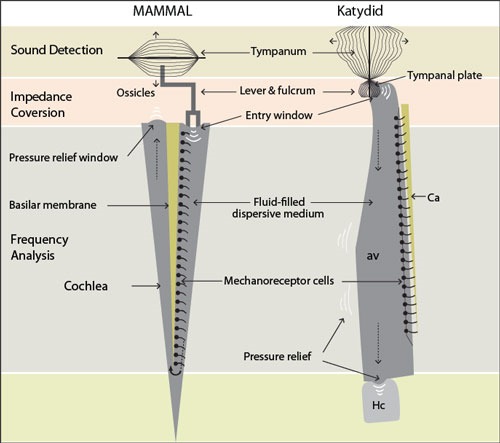
Figure 5 - Dorsolateral view of external ear and tarsus / CC-BY-SA-c.0
Dr. Fernando Montealegre-Zapata says that the “efficiency of this tiny system could inspire engineers to create microsensors based on the katydid’s ear design.” 11 Ron Hoy, a professor of neurobiology and behavior at Cornell University, wonders what katydid-inspired sensors “could be in the hands of an imaginative engineer?” 12
While the katydid’s hearing range is impressive, it doesn’t come close to the Greater Wax Moth, which can hear from 20,000 hertz to the record-breaking 300,000 Hz (Table 1). 13
|
|
Minimum Hz |
Maximum Hz |
|
Orange-Faced South American Katydid |
10,000 |
50,000 |
|
Gypsy Moth |
|
150,000 |
|
Greater Wax Moth |
20,000 |
300,000 |
|
Drosophila (Fruit Fly) |
100 |
1,000 |
|
Bats |
|
212,000 |
|
Humans |
20 |
20,000 |
The lead investigator of the research team that discovered the moth’s impressive hearing range, James Windmill, now wants to investigate how the moth manages to hear across such a wide frequency range, “As an engineer, I know that we would struggle to design an artificial version of this ear.” 14
These evolutionist scientists recognize the complexity and innovative design necessary for hearing, yet they repeatedly give the glory to evolution rather than the Creator. For auditory systems that are as complex as the ear to have occurred randomly multiple times, in multiple variations, defies all odds. Whatever the design of the auditory system, it seems to be perfectly suited to the creature. Each creature is given the auditory range needed to perform the functions that it was designed to fulfill. Fruit flies would not have survived if they had to evolve an antenna to hear their mate before they could mate. If they were already surviving without this antenna, an antenna tuned specifically to their possible mate’s wing frequency would definitely be an advantage, but what is the possibility that this would have occurred by random chance. Echolocation in bats and dolphins is only further evidence of the diverse creativity and imagination of our Creator. Yet this is only a small sampling of the variety of auditory systems. In part 2 I will share more evidence of the miracle of hearing.
- 1M. J (2012 Jul 25) On the evolution of the mammalian middle ear. https://evolutionnews.org/2012/07/on_the_evolutio/ Accessed 2017 Nov 30
- 2Parker J, Tsagkogeorga G, Cotton JA, Liu Y, Provero P, Stupka E, Rossiter SJ (2013) Genome-wide signatures of convergent evolution in echolocating mammals. Nature 502:228-31. http://www.nature.com/articles/nature12511 Accessed 2017 Nov 30
- 3Leweingast (2016 May 16) The survival of hearing: How physics has pressured genetic selection. https://pages.vassar.edu/sensoryecology/the-survival-of-hearing-how-physics-has-pressured-genetic-selection/ Accessed 2017 Nov 30
- 4De Beer G (1971) Homology: An Unsolved Problem. Carolina Biological Supply, Burlington
- 5Yager DD (1999) Structure, development, and evolution of insect auditory systems. Microsc Res Tech 47(6):380-400
- 6What-When-How (no author provided) Hearing (insects). http://what-when-how.com/insects/hearing-insects/ Accessed 2017 Nov 30
- 7Riabinina O, Dai M, Duke T, Albert JT (2011 Mar 31) Active process mediates species-species tuning of Drosophila ears. http://www.cell.com/current-biology/fulltext/S0960-9822(11)00273-9 Accessed 2017 Dec 02
- 8The Scientist staff (2017 Mar 1) Song around the animal kingdom. https://www.the-scientist.com/?articles.view/articleNo/48596/title/Song-Around-the-Animal-Kingdom/ Accessed 2017 Dec 02
- 9Albert JT, Gopfert MC (2015) Hearing in Drosophila. Curr Opin Neurobio 34:79-85 http://www.sciencedirect.com/science/article/pii/S0959438815000318 Accessed 2017 Dec 02
- 10Sarfati J (2013) Katydid’s amazing ear design. Creation 35(4): 12-13 https://creation.com/katydid Accessed 2017 Dec 02
- 11Sarfati J (2013) Katydid’s amazing ear design. Creation 35(4): 12-13 https://creation.com/katydid Accessed 2017 Dec 02
- 12Hoy RR (2012) Evolution. Convergent evolution of hearing. Science 338(6109):894–5
- 13Moir HM, Jackson JC, Windmill JFC (2013) Extremely high frequency sensitivity in a “simple” ear. Biol Lett 9:20130241 http://rsbl.royalsocietypublishing.org/ content/roybiolett/9/4/20130241.full.pdf Accessed 2017 Dec 02
- 14Yong E (2013 May 08) Moth smashes ultrasound hearing records. Nature https://www.nature.com/news/moth-smashes-ultrasound-hearing-records-1.12941 Accessed 2017 Dec 3